![]()
![]()
L'immunité peut être définie comme l'ensemble des mécanismes biologiques permettant à un organisme de reconnaître et de tolérer ce qui lui appartient (le soi) et de reconnaître et de rejeter ce qui lui est étranger (le non soi) : les substances étrangères ou les agents infectieux auxquels il est exposé, mais aussi ses propres constituants altérés (comme des cellules tumorales). La réponse immunitaire est déclenchée suite à la pénétration d'un agent étranger dans l'organisme et elle doit s'arrêter après l'élimination de cet agent étranger.
Le "Soi" et le "Non Soi"
La transfusion sanguine
Le système ABO
Pour préciser comment l'organisme est capable de reconnaître le "SOI" et le "NON SOI", on mélange des gouttes de sang de trois sujets A, B et C.
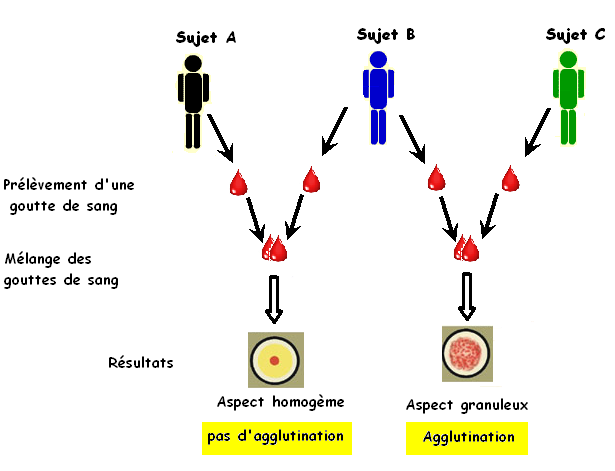
Le sang du sujet A et celui du sujet B sont compatibles, ona donc une tolérance d'ou l'absence d'une agglutination par contre le sang du sujet B et celui du sujet C sont incompatibles, on a une intolérance d'ou l'agglutination. L'agglutination est une réponse immunitaire qui confirme la présence du "NON SOI"
Pour préciser cette agglutination, on a mélangé des hématies et des plasmas des ces trois sujets A, B et C
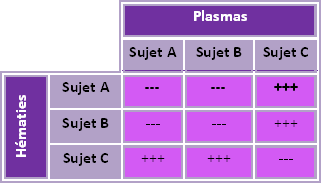
L'agglutination est une réaction spécifique entre les hématies et des molécules contenues dans le plasma
On a montré la présence des protéines marqueurs du SOI à la surface membranaire des hématies, il s'agit des glycoprotéines appelés des agglutinogènes. Il existe deux types des agglutinogènes: Les agglutinogènes A et les agglutinogènes B. On distingue quatre types des hématies:
Des hématies A: qui présentent uniquement les agglutinogènes A
Des hématies B: Qui présentent uniquement les agglutinogènes B
Des hématies AB: Qui présentent les agglutinogènes A et les agglutinogènes B
Des hématies O: qui ne présentent pas d'agglutinogènes (ni A ni B)
Dans le plasma on a trouvé des molécules spécifiques capables de réagir avec les agglutinogènes A et B, il s'agit des agglutinines ou des anticorps: les anti-A et les anti-B. On distingue quatre types de plasma:
Plasma qui contient uniquement des agglutinines anti-A
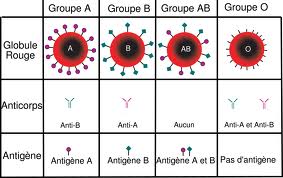
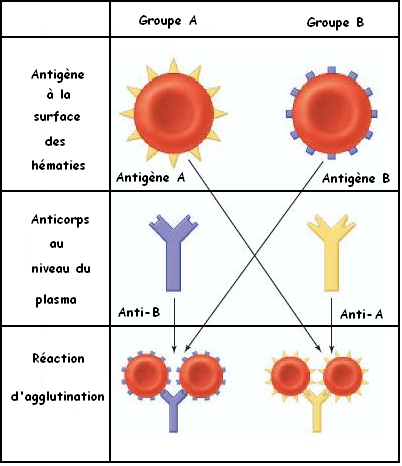
Pour déterminer les groupes sanguins des individus, on utilise les sérums test anti-A et anti-B
| Groupes Sanguins | anti-A Anti-B |
| Groupe O | |
| Groupe A | |
| Groupe B | |
| Groupe AB |
Le système ABO est contrôlé par un gène autosomal triallèlique (A,B,O) tel que
Les individus qui présentent un groupe sanguin positif possèdent un antigène marqueur du soi à la surface membranaire de leurs hématies: l'antigène Rh+. ceux qui ont un groupe sanguin négatif ne possèdent pas l'antigène Rh+
Les individus [Rh-] synthètisent des anticorps anti-Rh+ si on leurs introduit les antigènes Rh+
Le facteur Rhésus est contrôlé paur un caractère héréditaire (Rh+, Rh-) tel que l'allèle Rh+ domine l'allèle Rh-
La maladie hémolytique du nouveau-né
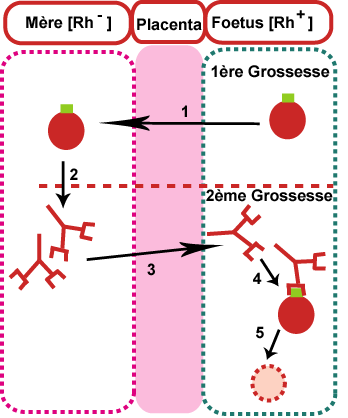
La maladie hémolytique du nouveau-né ne peut infecter que les enfants [Rh+] issus d'une mère [Rh-]. L'enfant atteint doit être précédé par la naissance d'un enfant [Rh+] sain
Au cours de la première grossesse, au moment de l'accouchement, quelques hématies du foetus [Rh+] peuvent passer vers la mère [Rh-] (1) qui déclenche une réponse immunitaire et synthétise des anticorps anti-Rh+ (2)
Au cours de la deuxième grossesse, les anticorps anti-Rh+ traversent le placenta (3) et passent vers le foetus [Rh+] provoquant une agglutination (4) et la lyse de ses hématies (5).
Les expériences de greffe et de transplantation
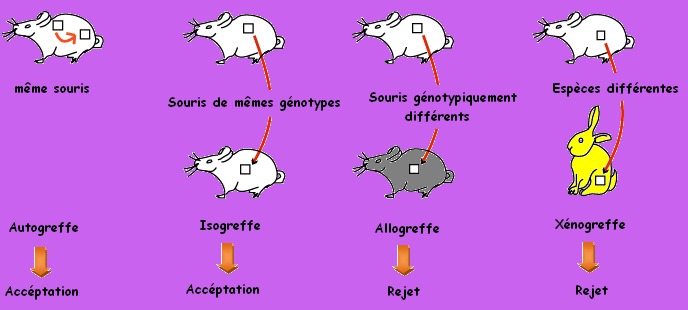
Le rejet du greffon obtenu dans les cas de l'allogeffe et le xénogreffe correspond à une réponse immunitaire qui confirme l'incompatibilité tissulaire entre les cellules greffées et les cellules du receveur.
Cette incompatibilité tissulaire est due à la présence des antigènes marqueurs du soi à la surface des toutes les cellules nucléées de l'organisme. ces antigènes marqueurs du soi sont codés par des gènes liés et polyalléliques qui forment un complexe appelé le Complexe Majeur d'Hitocompatibilité (CMH). Chez l'espèce Humaine le CMH s'appelle le HLA. Chaque individu hérite deux allèles de chacun des ces gènes et dispose d'une combinaison allèlique originale. Les individus qui présentent des combinaison allèliques différentes, présentent des antigènes marqueurs du soi différents donc des HLA ou CMH différents.
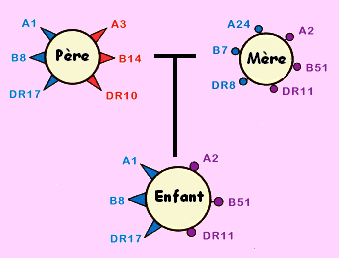
On distingue deux classes de HLA ou CMH
HLA ou CMH I: à la surface de toutes les cellules nucléées et qui intervient dans le rejet de greffe
HLA ou CMH II: à la surface de cellules immunitaires: lymphocytes et macrophage
Les propriétés de la réponse immunitaire spécifique
L'acquisition
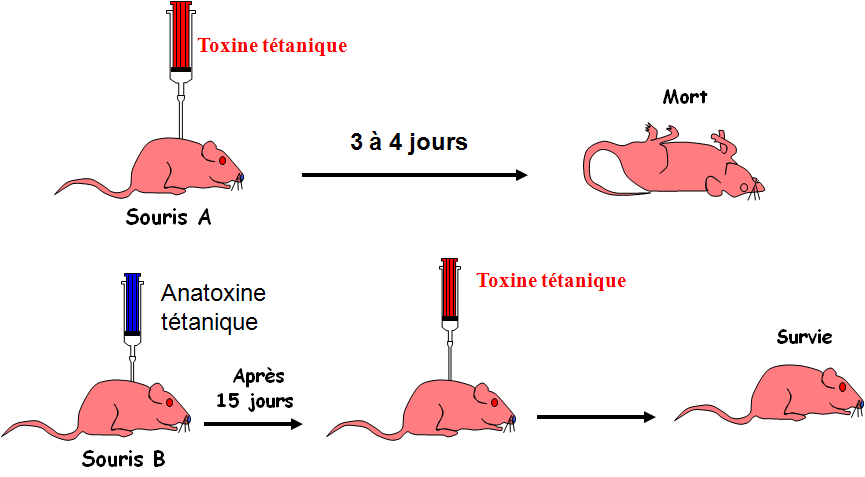
La spécificité
La protection acquise contre le tétanos ne protège pas la souris C contre la diphtérie donc la réponse immunitaire est spécifique.
La mémoire
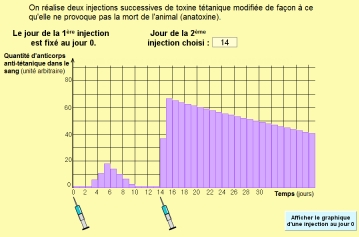
Des lymphocytes ayant eu un 1er contact avec un antigène sont toujours conservés : on parle de lymphocytes mémoire.
Si un 2ème contact a lieu avec le même antigène, ils se multiplient plus vite, produisent plus d’anticorps. La réponse immunitaire secondaire est plus forte et plus rapide que la réponse primaire.
La vaccination utilise cette propriété :
- on présente à l’organisme des microbes atténués ou des antigènes sans pouvoir pathogène (anatoxines)
- on présente une 2ème fois les antigènes aux lymphocytes, afin de renforcer la réponse immunitaire
- en cas de contamination ultérieure, le système immunitaire est prêt à répondre très fort et très vite.
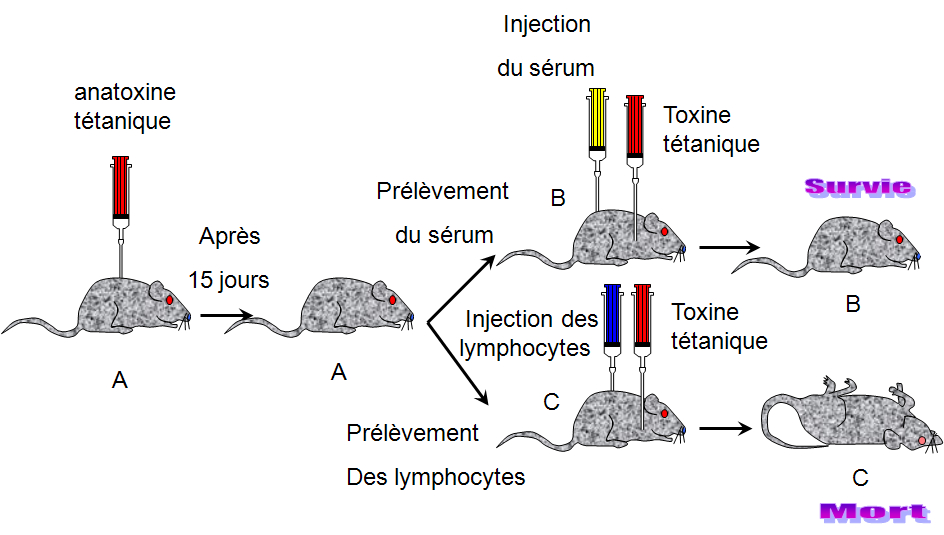
La transférabilitéé
![]()
La souris B est protégée par le sérum de la souris A immunisée contre le tétanos, donc on a un transfert de l'immunité de la souris A immunisée à la souris B non immunisée.
La diversité
L'injection d'un antigène X déclenche une réponse immunitaire par la synthèse des anticorps anti-X, l'injection d'un antigène Y, déclenche une réponse immunitaire par la synthèse des anticorps anti-Y..... les anticorps synthétisés sont différents puisque les antigènes sont différents et puisque la réponse immunitaire est spécifique. donc la réponse immunitaire est caractérisée par une diversité.
Les voies de la réponses immunitaire spécifique
La RIMH
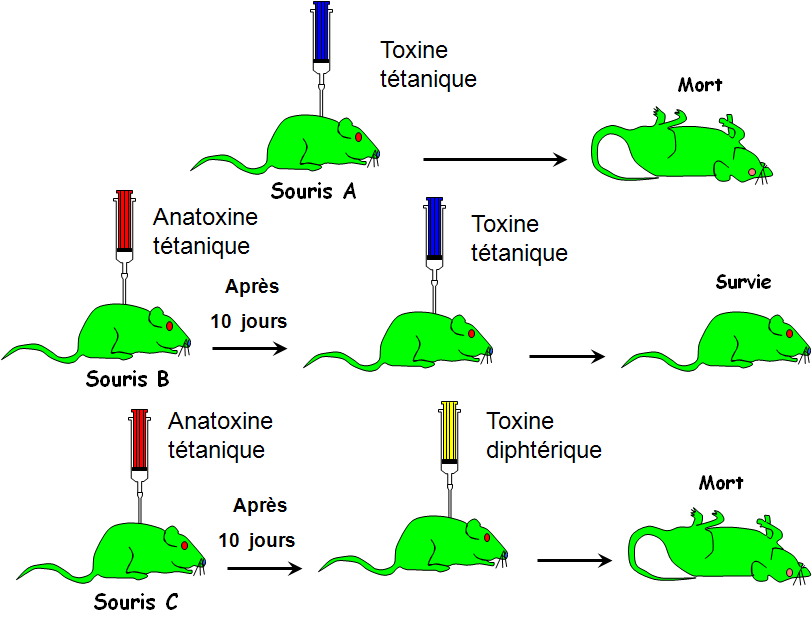
Le sérum de la souris A immunisée contre le tétanos protège la souris B contre la toxine tétanique par contre les lymphocytes de la souris A sont incapables de protéger la souris C contre la toxine tétanique: On a un transfert de l'immunité de la souris A à la souris B par l'intermédiaire du sérum qui contient des anticorps il s'agit donc d'une réponse immunitare à médiation humorale : RIMH
La RIMC
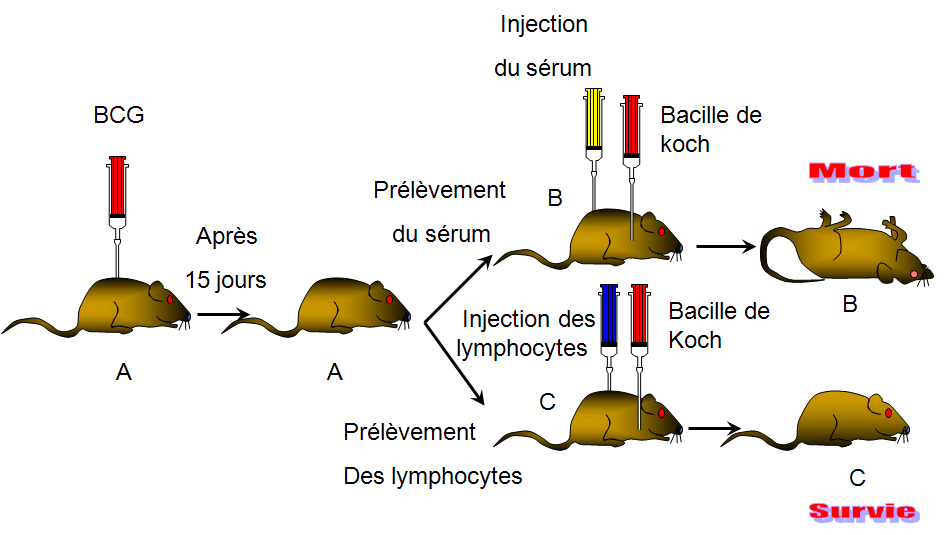
Les lymphocytes de la souris A immunisée contre la tuberculose protège la souris C contre la bacille de Koch par contre le sérum de la souris A est incapable de protèger la souris B contre la bacille de Koch: On a un transfert de l'immunité de la souris A à la souris C par l'intermédiaire des lymphocytes (cellules) il s'agit d'une réponse immunitaire à médiation cellulaire: RIMC
Les organes et les cellules lymphoïdes
La réponse immunitaire spécifique est assurée par des cellules immunitaires: des leucocytes (des globules blancs) on distingues des lymphocytes, des monocytes qui se tranforment en macrophages et des granulocytes qui se transforment en polynucléaires.toutes les cellules immunitaires sont nées au niveau de la moelle osseuse rouge
| Naissance | Maturation | |
|
Lymphocytes B |
Moelle osseuse rouge | Moelle osseuse rouge |
| Lymphocytes T | moelle osseuse rouge |
Thymus |
Les lymphocytes B (LB) et les lymphocytes T (LT) sont nés au niveau de la moelle osseuse rouge à partir des cellules souches lymphoïdes. La cellule souche produit des lymphocytes non immunocompétents des pré-B et des pré-T incapables de reconnaitre l'antigène et de déclencher une réponse immunitaire. Les pré-B restent dans la moelle osseuse et complètent leur maturation por donner des LB immunocompétents par contre les pré-T migrent vers le thymus où ils terminent leur maturation pour donner des LT immunocompétent. Au niveau du thymus se forment deux catégories des LT: des LT4 et des LT8. Les LB et les LT (LT8 et LT4) quittent respectivement la moelle osseuse et le thymus pour aller vers les organes lymphoïdes secondaires (les ganglions lymphatiques, la rate et les amygdales) où ils peuvent rencontrer les antigènes.
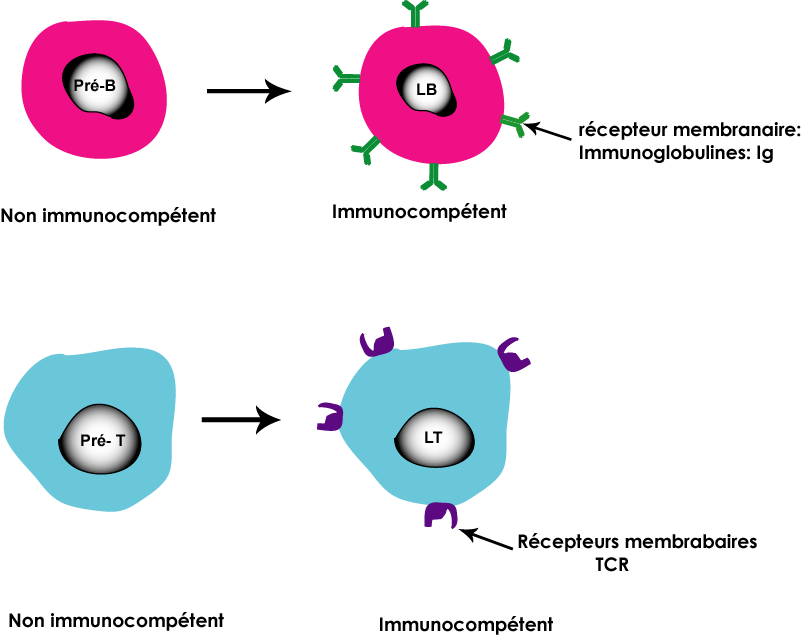 La maturation des lymphocytes LB et LT consiste à une acquisition de l'immunocompétence grâce à la synthèse des récepteurs membranaires
La maturation des lymphocytes LB et LT consiste à une acquisition de l'immunocompétence grâce à la synthèse des récepteurs membranaires
La synthèse des immunoglobulines (Ig) pour les LB
La synthèse des TCR pour les LT (LT8 et LT4)
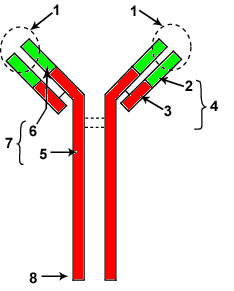 1-site d'anticorps ou site de fixation de l'antigène
1-site d'anticorps ou site de fixation de l'antigène
2- zone variable de la chaine légère (VL)
3- zone constante de la chaine légère (CL)
4- chaine légère (L)
5-zone constante de la chaine lourde (VH)
6-zone variable de la chaine lourde (CH)
7-chaine lourde (H)
8-site effecteur
Les immunoglobulines (Ig) sont des glycoprptéines formées de quatre chaines identiques deux à deux deux chaines légères (L) et deux chaines lourdes (H) Chaque LB ne peut synthétiser qu'un seul type de Ig et par conséquent ne peut reconnaitre qu'un seul type d'antigène (un seul épitope ou déterminant antigénique) On dispose donc d'un répertoire complet des LB différents par leurs Ig.

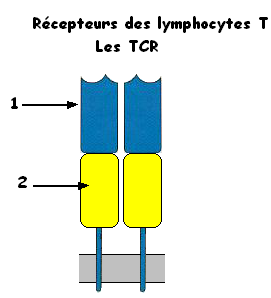
1- zone variable
2- zone constante
Les TCR sont formés de deux chaines identiques. les extrémités des zones variables représentent le site de reconnaissance de l'antigène qui présente deux sites de fixation: un site de fixation de l'antigène (épitope) et un site pour le CMH (HLA)
Le déroulement de la réponse Immunitaire
La phase d'induction
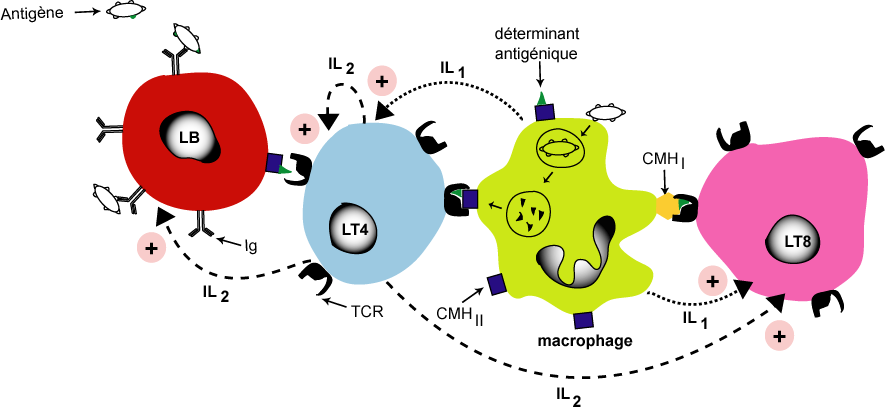
Cette phase est caractérisée par la reconnaissance de l'antigène, la coopération entre les cellules immunitaires (LB, LT et macrophage) et l'activation des lymphocytes (LB, LT4 et LT8)
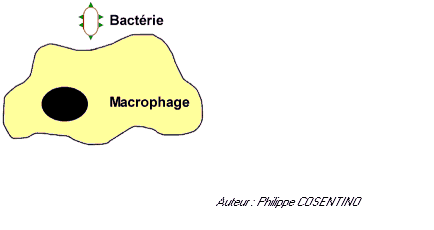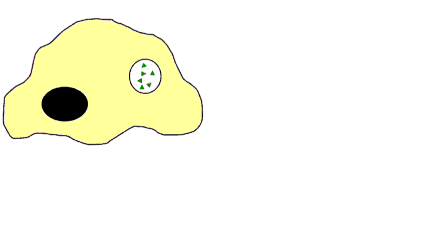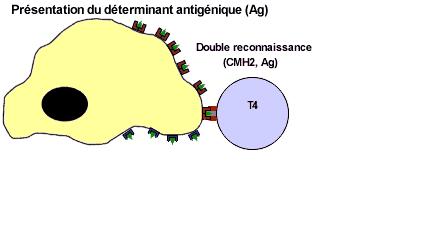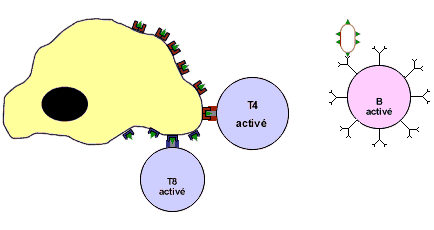
La reconnaissance de l'antigène
![]() Reconnaissance directe et simple de l'antigène pour les macrophages et les LB. ces cellules sont capables de reconnaitre des antigènes libres grâce à des récepteurs membranaires
Reconnaissance directe et simple de l'antigène pour les macrophages et les LB. ces cellules sont capables de reconnaitre des antigènes libres grâce à des récepteurs membranaires
![]() Recnnaissance indirecte et double de l'antigène pour les LT4 et les LT8. Ces cellules reconnaissent l'antigène associé au CMHI ou au CMHII grâce à leurs récepters membranaires les TCR. Les LT4 reconnaissent l'antigène associé au CMHII par contre les LT8 reconnaissent l'antigène associé au CMHI. Les LT4 et les LT8 nécessite une cellule présentatrice de l'antigène: une CPAg (macrophage ou LB)
Recnnaissance indirecte et double de l'antigène pour les LT4 et les LT8. Ces cellules reconnaissent l'antigène associé au CMHI ou au CMHII grâce à leurs récepters membranaires les TCR. Les LT4 reconnaissent l'antigène associé au CMHII par contre les LT8 reconnaissent l'antigène associé au CMHI. Les LT4 et les LT8 nécessite une cellule présentatrice de l'antigène: une CPAg (macrophage ou LB)
La coopération cellulaire et l'activation des lymphocytes
La RIMH et la RIMC nécessitent une coopération cellulaire entre les macrophages, les LB, et les LT.
![]() Une coopération directe par contact direct entre les lymphocytes et le macrophage: entre macrophage et LT8, entre macrophage et LT4 et entre LB et LT4
Une coopération directe par contact direct entre les lymphocytes et le macrophage: entre macrophage et LT8, entre macrophage et LT4 et entre LB et LT4
![]() Une coopération indirecte par l'intermédiaire des substances chimiques appelées les interleukines (IL1 et IL2)
Une coopération indirecte par l'intermédiaire des substances chimiques appelées les interleukines (IL1 et IL2)
![]() L'interleukine (IL1) est sécrété par le macrophage pour activer les LT4 et les LT8
L'interleukine (IL1) est sécrété par le macrophage pour activer les LT4 et les LT8
![]() L'interleukine (IL2) est sécrété par les LT4 activés par IL1, pour activer Les LT8 et Les LB et pour une auto-activation (activation de LT4)
L'interleukine (IL2) est sécrété par les LT4 activés par IL1, pour activer Les LT8 et Les LB et pour une auto-activation (activation de LT4)
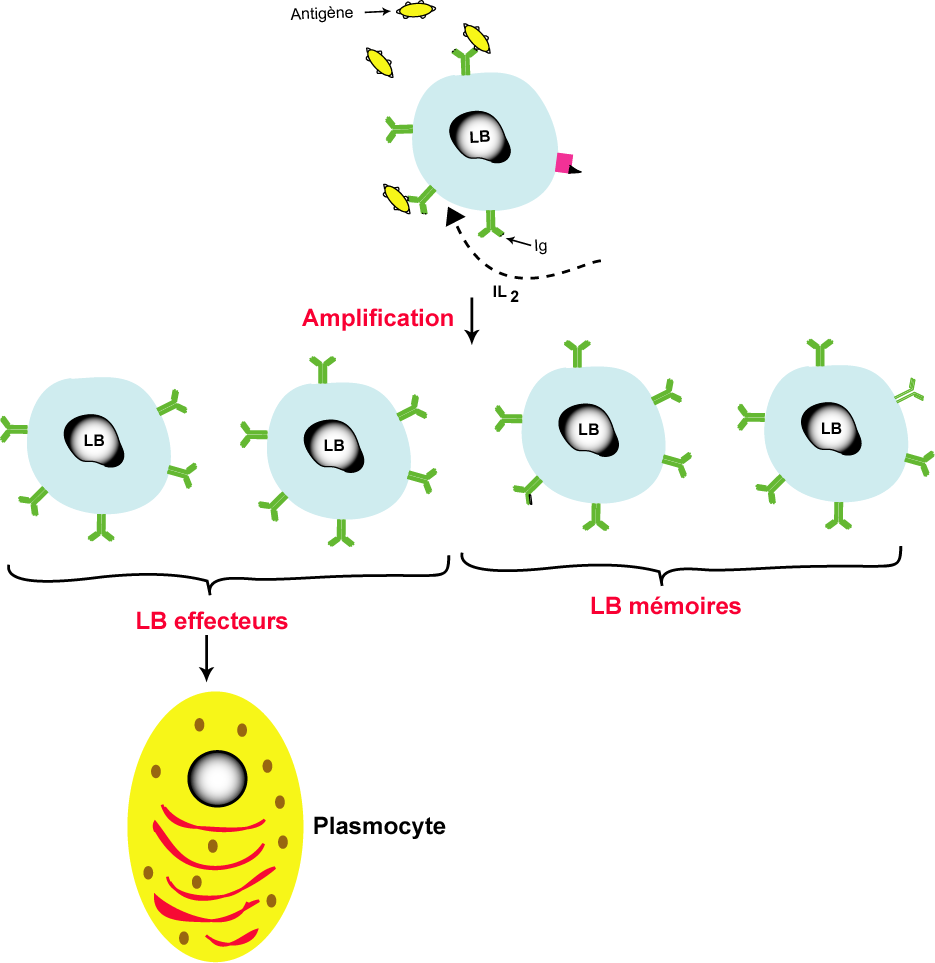
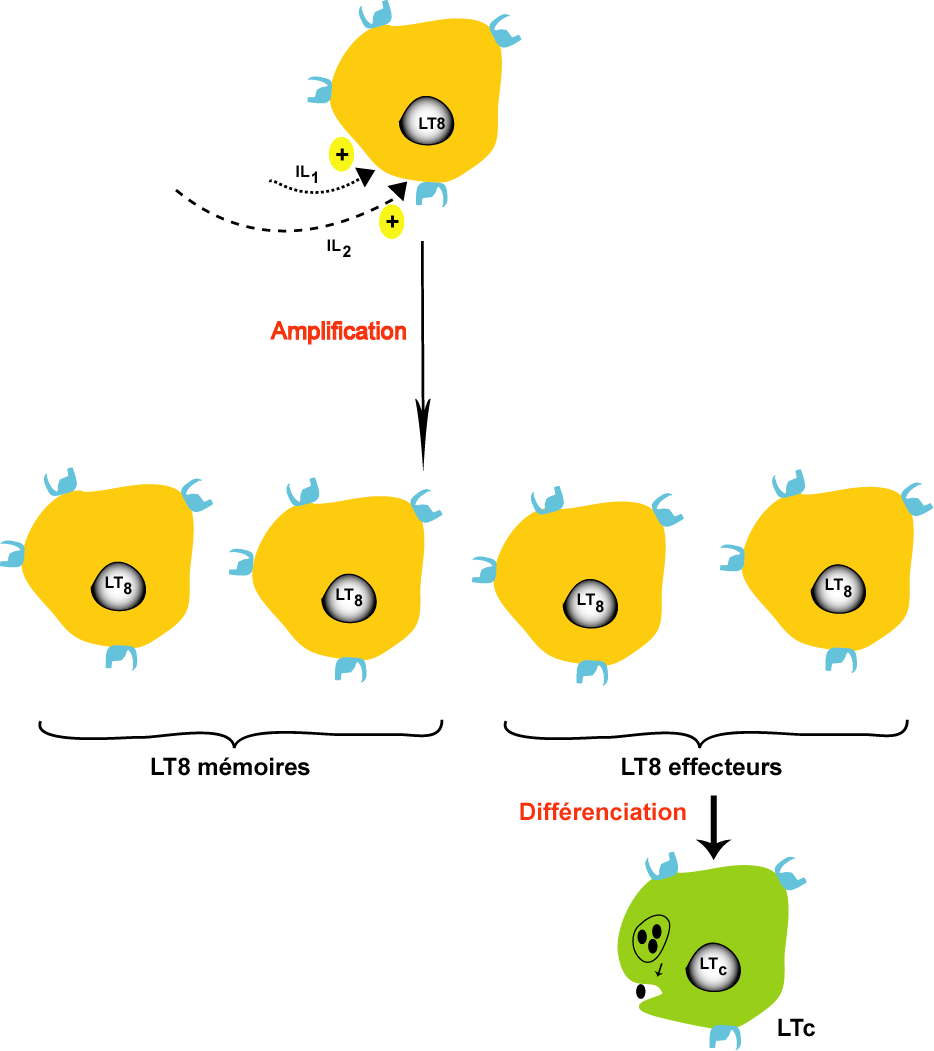
La phase d'amplification et de différenciation
Les lymphocytes activés (LB, LT4 et LT8) subissent une multiplication active par des mitoses successives pour donner des clones qui comportes de lymphocytes mémoires et des lymphocytes effecteurs. les lymphocytes mémoires interviennent au cours de la réponse immunitaire secondaire par contre les lymphocytes effeteurs subissent une différenciation pour donner des cellules différenciées
Les LB effecteurs se différencient en plasmocytes sécrétrices d'anticorps libres (anticorps sériques)
Les LT4 effecteurs se différencient en LT auxiliaires (LTa) encore appelés LT helper (LTh) sécrétrices d'interleukine 2 (IL2)
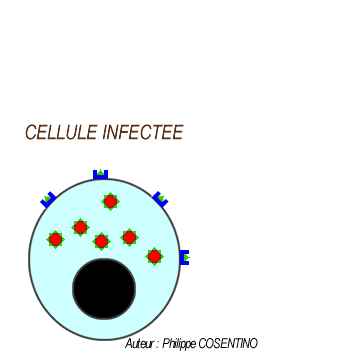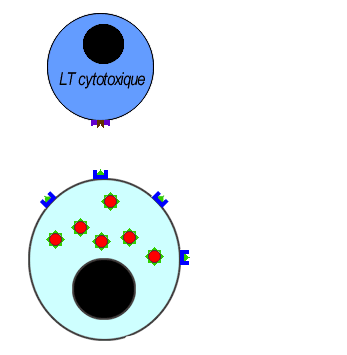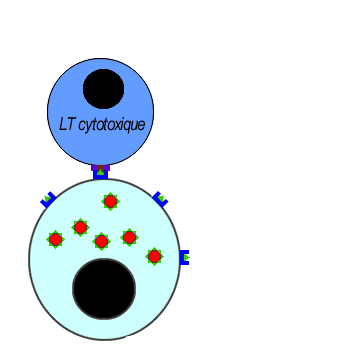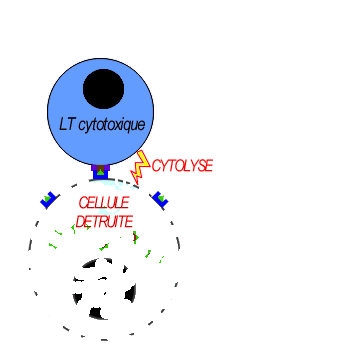
Les LT8 effecteurs se différencient en LT cytotoxiques (LTC) sécrétrices de perforines pour assurer la lyse des cellules infectées par l'antigène.
un groupe de LT8 effecteurs se différencient en LT suppresseurs (LTs) qui interviennent après l'élimination de l'antigène pour supprimer la réponse immunitaire.

La phase effectrice
![]() Dans le cas de la RIMH
Dans le cas de la RIMH
Les anticorps libres synthéthisés et sécrétés par les plasmocytes réagissent avec l'antigène pour former un complexe immun: on a une neutralisation de l'antigène. Le complexe immun est par la suite détruit par
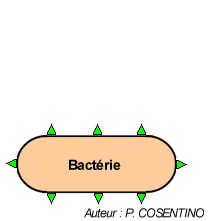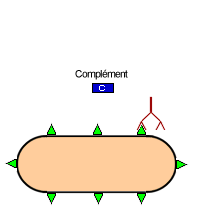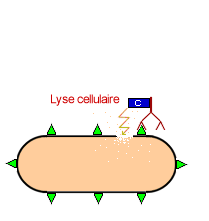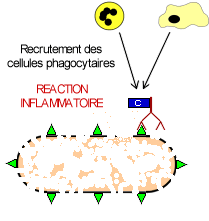
![]() Le complément: ensemble des molécules (enzymes) qui se fixent sur le complexe immun et provoque sa lyse
Le complément: ensemble des molécules (enzymes) qui se fixent sur le complexe immun et provoque sa lyse
![]() Opsonisation: le complexe immun est phagocyté par une cellule phagocytaire (macrophage ou polynucléaire)
Opsonisation: le complexe immun est phagocyté par une cellule phagocytaire (macrophage ou polynucléaire)
![]() Dans le cas de la RIMC
Dans le cas de la RIMC
Les lymphocytes LTC reconnaisent l'antigène associé au HLAI grâce à leurs récepteurs (le TCR) cette fixation déclenche la libération des perforines et des enzymes. les molécules de perforine se polymérisent sur la membrane des cellules infectées créant des canaux favorisant l'entrée d'eau et des enzymes hydrolytiques provoquant la lyse des cellules infectées.